PB Ch 36. Distant Hybridization
1.1 Introduction and Definition
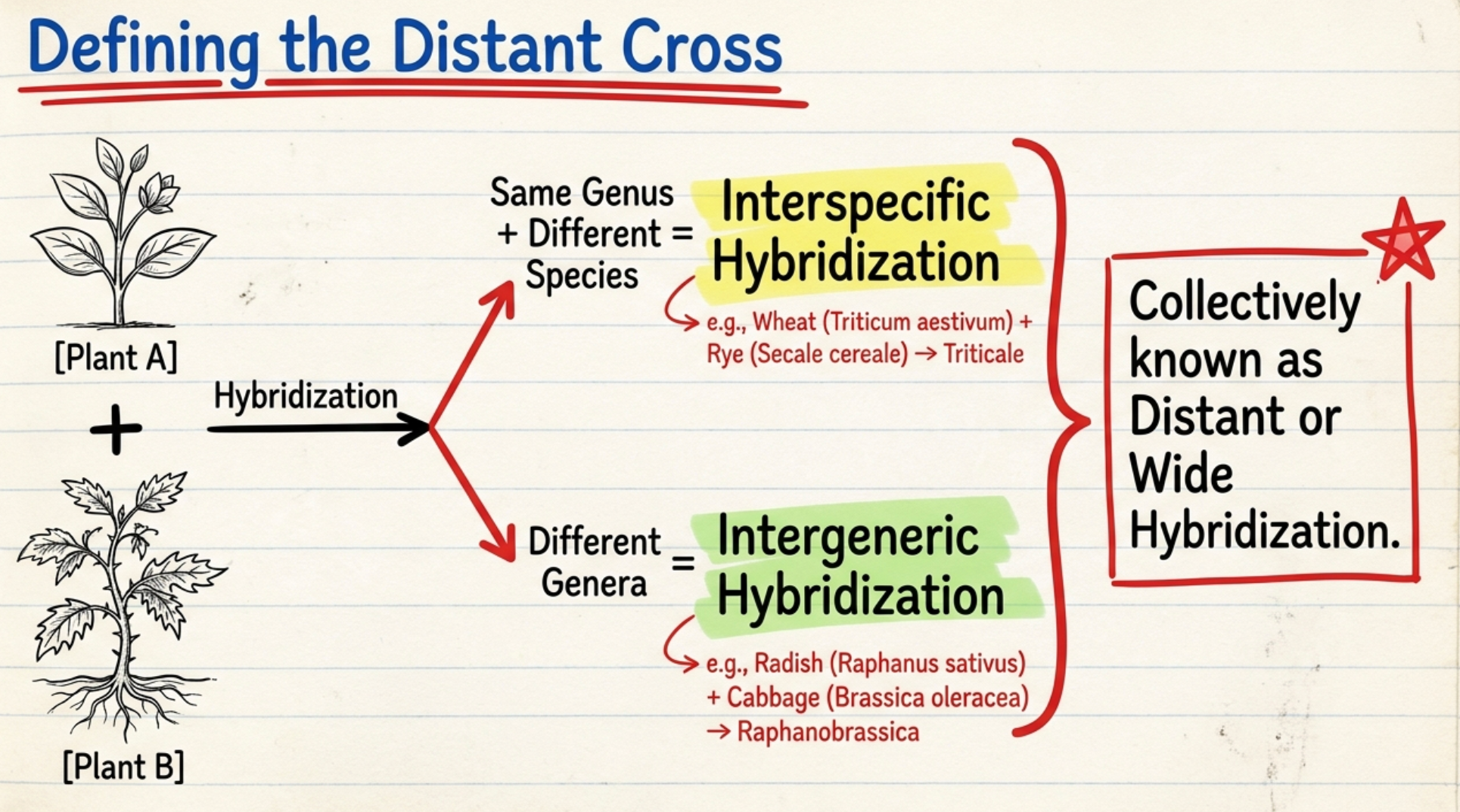
- When hybridization is performed between individuals from different species of the same genus, it is known as interspecific hybridization.
- When the individuals being crossed belong to two different genera, it is referred to as intergeneric hybridization.
- Hybridization between individuals from different species of the same genus, or two different genera, is collectively termed distant hybridization or wide hybridization, and such crosses are called distant crosses or wide crosses.
- Distant hybridization is the principal method used to access genes present in wild relatives and related species that are not available in the cultivated gene pool. It has produced some of the most important crop improvements in the 20th century — particularly in disease resistance and the creation of entirely new crop species.
1.2 History of Distant Hybridization
- 1717: Thomas Fairchild — first recorded distant hybrid: carnation (Dianthus caryophyllus) × sweet William (Dianthus barbatus) — called Fairchild's Mule. This is also the first documented artificial plant hybrid.
- Early interspecific hybrids: Most early interspecific hybrids were of no agricultural value. However, many interspecific hybrids in ornamentals served as commercial varieties.
- 1928: Karpechenko — Raphanobrassica, an interesting amphidiploid between radish (Raphanus sativus) and cabbage (Brassica oleracea). This was the first experimentally produced allopolyploid and demonstrated that hybridization between species of different genera could produce a fertile new species, but the hybrid was agronomically useless — it had the root of a cabbage and the leaves of a radish.
- 1890: Rimpau — first Triticale hybrid, the first intergeneric hybrid of great agricultural potential. Modern Triticale development occurred at CIMMYT (I believe this is approximately correct — please verify the exact institutional history from CIMMYT publications).
- 20th century: Rapid expansion of distant hybridization for disease resistance, particularly in wheat (rust resistance from Aegilops), rice (grassy stunt resistance from O. nivara), and cotton (fibre quality from G. barbadense).
1.3 Objectives of Wide Hybridization
1. Transfer of Desirable Characters from Wild Relatives
- Disease resistance: Many disease resistance genes not available in cultivated species exist in wild relatives. Examples: many of the genes for rust resistance in wheat (Lr genes for leaf rust, Sr genes for stem rust, Yr genes for yellow rust) are derived from related wild Aegilops and Triticum species. TMV resistance in tobacco from Nicotiana glutinosa.
- Insect resistance: Wild relatives of cotton, rice, and tobacco carry genes for resistance to sucking insects and bollworm not found in cultivated varieties.
- Wide adaptability: Cold tolerance has been transferred from wild relatives to wheat, onion, potato, tomato, and grape.
- Quality improvement: Oil quality in oil palm was improved by genes from wild relatives. Fibre length in Indian cotton (Gossypium arboreum) was improved by crossing with American cotton.
- Yield improvement: Distant hybridization has contributed to yield improvement in oats, tobacco, maize, and sugarcane.
- Cytoplasmic male sterility (CMS): CMS sources used in commercial hybrid seed production in sorghum, sunflower, and other crops were derived from interspecific hybridization with wild relatives.
2. Exploitation of Luxuriance (Heterosis)
- In vegetatively propagated crops and ornamentals, the extremely high heterosis (luxuriance) observed in interspecific hybrids can be exploited directly as F1 hybrid varieties, since the hybrid genotype can be maintained clonally.
- Examples: sugarcane varieties, ornamental chrysanthemum.
3. Creation of Novel Genotypes
- Distant hybridization followed by chromosome doubling can create entirely new allopolyploid species not previously existing in nature.
- Triticale (wheat × rye) is the most successful example. Raphanobrassica (radish × cabbage) is a scientific curiosity.
- Brassica napus (canola/oilseed rape) is another allopolyploid species of great agricultural importance.
1.4 Barriers to Production of Distant Hybrids
Distant hybridization faces several biological barriers. These are classified into three main categories:
Category 1: Failure of Zygote Formation (Cross-incompatibility / Pre-fertilization Barriers)
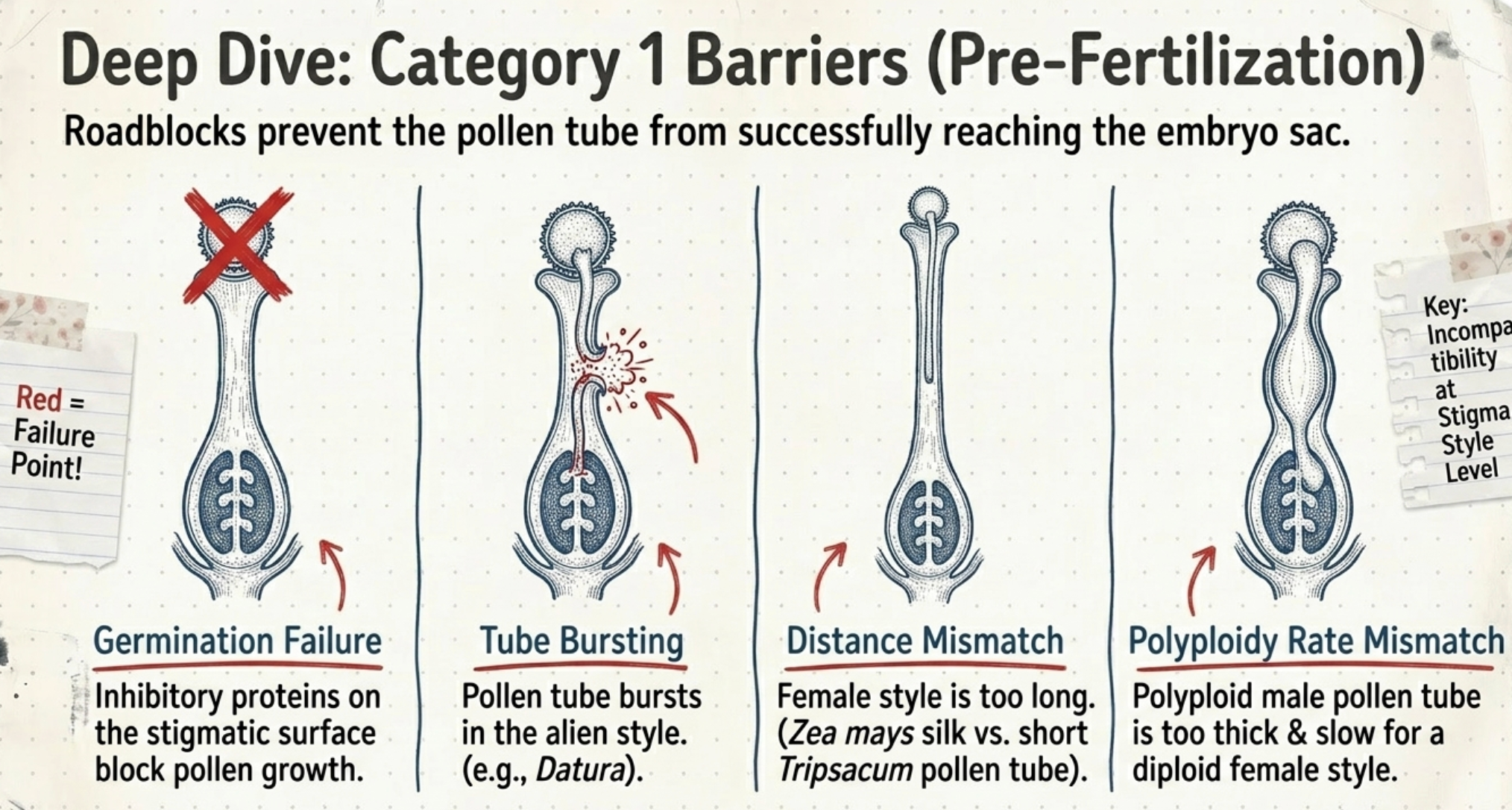
Inability of functional pollen of one species to effect fertilization of female gametes of another species. This is caused by:
- Pollen fails to germinate: On an alien stigma, the pollen may fail to germinate due to inhibitory proteins on the stigmatic surface.
- Pollen tube unable to reach embryo sac: The pollen tube may grow down the style but fail to reach the embryo sac due to style-pollen tube incompatibility
- Pollen tube bursts in the style: In Datura, pollen tube bursts within the style of a different species.
- Style longer than pollen tube growth: When the style of the female is longer than the normal growth of the male pollen tube, fertilization is prevented. Example: Zea mays × Tripsacum sp. — maize silk is much longer than the pollen tube of Tripsacum can traverse.
- Polyploidy pollen tube rate: Pollen tubes of polyploid species are usually thicker. When a diploid is used as female and a polyploid as male, the polyploid pollen tube grows at a slower rate in the diploid style than it would in a polyploid style, preventing timely fertilization.
Category 2: Failure of Zygote Development (Hybrid Inviability / Post-fertilization Barriers)
Fertilization occurs but the hybrid embryo fails to develop under normal conditions. Causes:
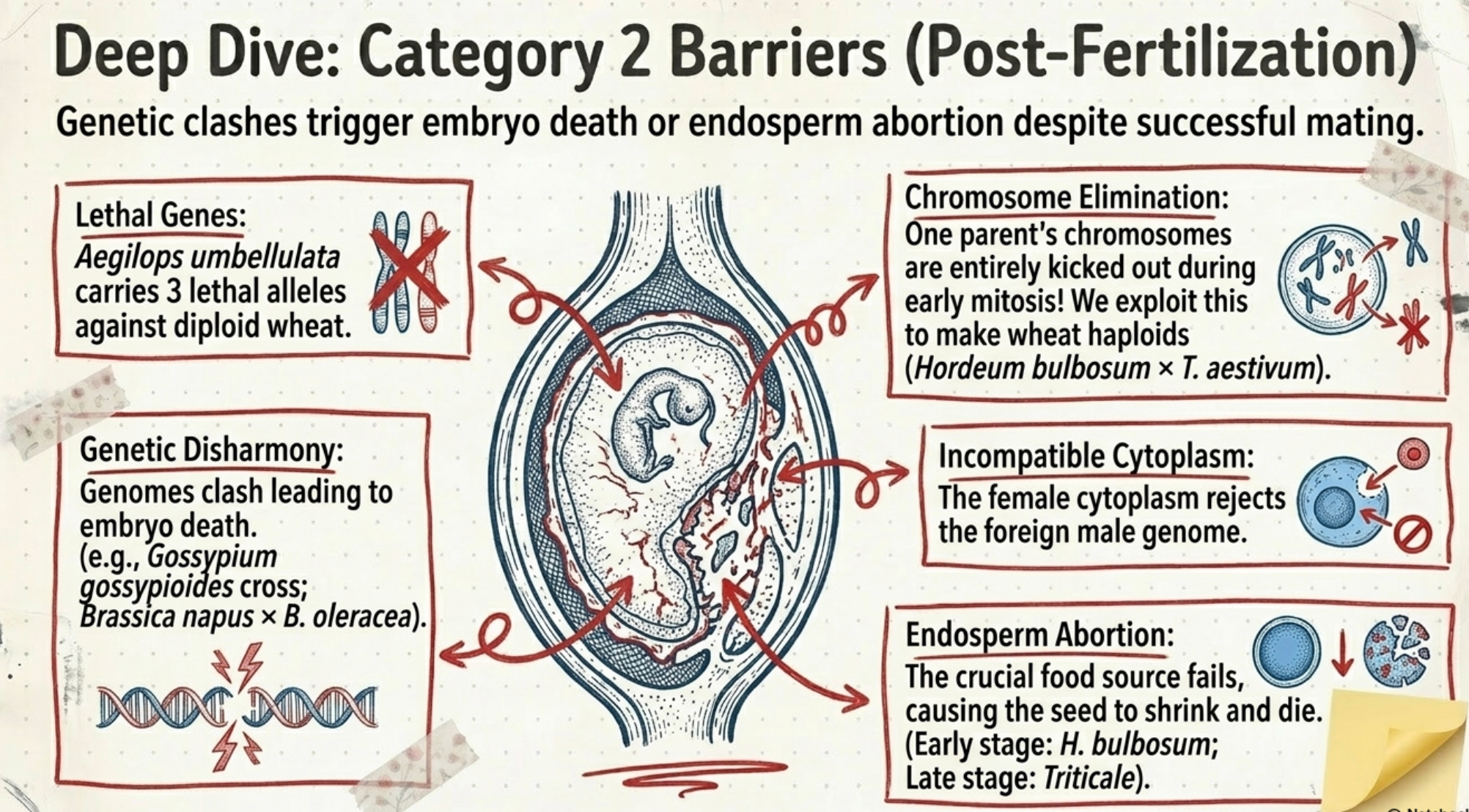
- Lethal genes: Some species carry lethal genes that cause death of the interspecific hybrid zygote during early embryonic development. Example: Aegilops umbellulata carries a lethal gene with three alleles against diploid wheats.
- Genetic disharmony: The genomes of the two parental species may be so different that their combined action leads to genetic imbalance, causing embryo death. Examples: Gossypium gossypioides × other Gossypium species; Brassica napus × B. oleracea.
- Chromosome elimination: In some distant crosses, chromosomes from one parent are gradually eliminated from the hybrid cells during early mitotic divisions of the zygote. The resulting embryo is not a true interspecific hybrid. Examples: Hordeum bulbosum × H. vulgare; Hordeum bulbosum × Triticum aestivum; Triticum aestivum × Zea mays. This chromosome elimination property has been exploited to produce wheat haploids (wheat × maize system).
- Incompatible cytoplasm: Incompatibility between the cytoplasm of the female parent and the genome of the male parent may block embryo development or lead to hybrid weakness and male sterility in the hybrid.
- Endosperm abortion: Seeds from many distant crosses are shrunken due to poorly developed endosperm (seeds show poor germination). When endosperm development is blocked, the condition is called endosperm abortion. The embryo can survive if rescued. Examples: Triticum × Secale (Triticale) — endosperm aborts at a later stage, giving a small frequency of viable seeds; Hordeum bulbosum × H. vulgare — endosperm aborts at an early stage, no viable seeds produced without embryo rescue.
Category 3: Failure of Hybrid Seedling Development / Hybrid Sterility
The F1 hybrid plants are produced but are completely or partially sterile, or seedlings die before maturity.
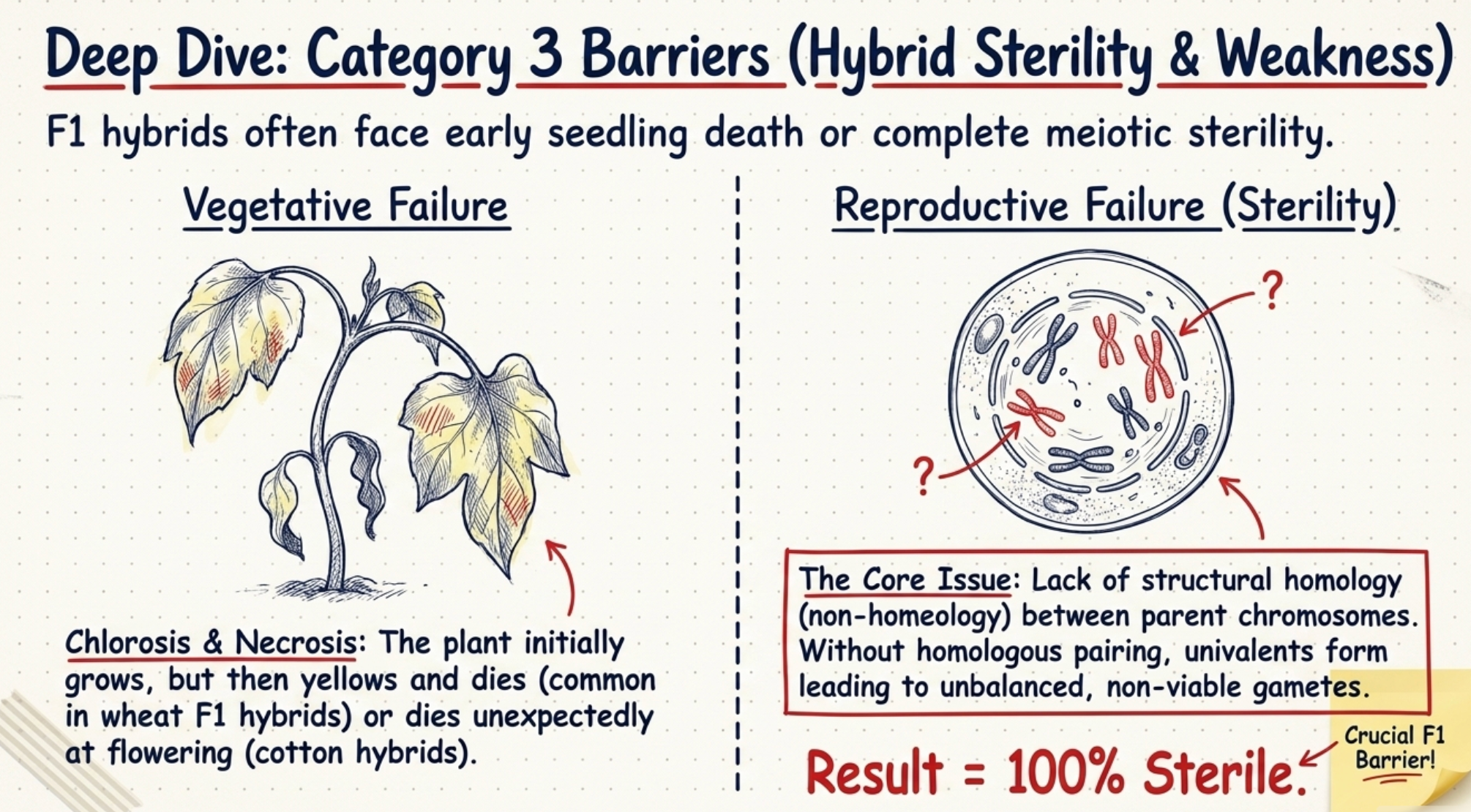
- Death during seedling development: Some interspecific hybrids appear normal initially but die at various stages of seedling growth; some plants die at flowering. Example: certain interspecific hybrids in cotton.
- Chlorosis and necrosis: Interspecific and intergeneric F1 hybrids of wheat show both chlorosis and necrosis, affecting plant development.
- Hybrid sterility: The main cause is lack of structural homology (non-homeology) between chromosomes of the two species. Without homologous pairing at meiosis, chromosomes form univalents, distributing irregularly into gametes and producing unbalanced, non-viable gametes.
|
1.5 Techniques to Overcome Barriers in Wide Hybridization
1. Choice of Parents
- Genetic differences exist among accessions within a species for cross-compatibility.
- Parents that show greater crossability should be selected for wide crosses.
- Testing multiple accessions of the wild species increases the chances of finding a compatible genotype.
- In wheat, for example, some accessions of Aegilops species cross more readily with cultivated wheats than others.
2. Pollination of Sufficiently Large Numbers of Flowers
- Success rate in wide crosses is generally very low.
- Pollinating a large number of flowers — hundreds to thousands — ensures that even a small percentage of successful fertilisations yields enough hybrid seeds to work with.
3. Reciprocal Crosses
- When a direct cross (A ♀ × B ♂) fails, the reciprocal cross (B ♀ × A ♂) should always be attempted.
- The direction of the cross matters in distant hybridization because the female parent's cytoplasm and stigmatic environment can be very different.
- Example: Phaseolus aureus and P. mungo are crossable only when P. aureus is used as female and P. mungo as male — not the reverse.
4. Manipulation of Ploidy Level
When two species differ in chromosome number or ploidy level, manipulating ploidy can increase crossability:
- Use higher ploidy species as female parent: A polyploid as female accepts pollen from a diploid more readily than a diploid female accepts polyploid pollen.
- Double chromosomes of wild species: Making the wild species autotetraploid before crossing with a tetraploid crop can bridge the ploidy barrier.
- Example: Brassica oleracea (CC, 2n=18) × B. campestris (AA, 2n=20) → B. napus (AACC, 2n=38). The two diploid parents have to be crossed as autotetraploids to produce the amphidiploid B. napus — this cross is very difficult without ploidy manipulation and embryo culture.
5. Bridge Crosses
- When two species A and C cannot be crossed directly, a third species B is chosen as a bridge. B must be crossable with both A and C. First B × C is made, then the F1 or amphidiploid from that cross is used as a bridge to cross with A.
- Tobacco example: Nicotiana repanda cannot be directly crossed with N. tabacum. However, N. sylvestris can cross with both N. repanda and N. tabacum. For gene transfer from N. repanda to N. tabacum, N. sylvestris serves as the bridge species: N. repanda × N. sylvestris → F1 or amphidiploid → crossed with N. tabacum.
- Wheat example: To transfer genes from Aegilops umbellulata (CC genome) to bread wheat (AABBDD), an intermediate amphidiploid can be used as a bridge.
6. Use of Pollen Mixtures (Mentor Pollen)
- Cross-incompatibility can result from unfavourable protein interactions between pistil and pollen.
- This can sometimes be overcome by mixing incompatible pollen (male × female pollen) with compatible pollen (same species as female, or irradiated killed pollen of same species).
- The compatible (mentor) pollen provides diffusible 'recognition' substances that allow the incompatible pollen to germinate and grow.
- Irradiated pollen from the same species is sometimes used as mentor to avoid self-fertilization.
7. Manipulation of Pistil (Stub Pollination)
- When pollen tube growth is limited and the style of the female is very long (preventing the pollen tube from reaching the ovule), the style can be cut to normal size before pollination.
- The technique is called stub pollination or cut-style pollination.
- This is successful in maize × Tripsacum crosses, where the maize silk remains receptive even after cutting.
8. Use of Growth Regulators
- In some cases, the pollen tube grows so slowly that the egg cell dies or the flower aborts before male gametes reach the ovary.
- Growth regulators can be used to accelerate pollen tube growth or prolong pistil viability. IAA, NAA, 2,4-D, and GA3 have shown promise in some wide crosses.
- Cytokinin and GA3 application to the ovary sometimes prevents flower/ovule abortion after fertilization.
9. Embryo Rescue (Embryo Culture)
- This is the most widely used technique to overcome post-fertilization barriers in distant hybridization.
- When the hybrid embryo aborts due to endosperm failure or incompatibility, the immature embryo is aseptically excised and cultured on an artificial nutrient medium before it aborts naturally.
Procedure:
- The wide cross is made and the female plant is allowed to develop fruits.
- Fruits are collected at 10–25 days after pollination (before natural abortion, but after the embryo has developed enough to be manipulated).
- Seeds are surface-sterilised using alcohol and sodium hypochlorite.
- Under sterile conditions (laminar flow cabinet), embryos are aseptically excised from the ovule.
- Embryos are placed on MS (Murashige and Skoog) medium supplemented with sucrose (approximately 3%), phytohormones (BAP for shoot, NAA for root), and agar.
- The embryo germinates and grows into a plantlet.
- Plantlets are hardened and transferred to the field.
- When the embryo is too small or inaccessible to excise, the entire ovule (ovule culture) or even the ovary (ovary culture) is cultured. Ovule culture is used in papaver, iris, and some Brassica crosses.
10. Chromosome Doubling with Colchicine
- Sterile F1 interspecific hybrids can often be made fertile by doubling their chromosome number using colchicine.
- Colchicine inhibits spindle fibre formation at metaphase, preventing cell division while allowing chromosome replication — resulting in cells with twice the chromosome number.
- The resulting amphidiploid has two complete chromosome sets (one from each parent species) and behaves as a diploid at meiosis — regular bivalent formation and normal fertility return.
- This technique (developed by Blackslee and Nebel in 1937) is the basis for all artificially produced allopolyploids — including Triticale, Raphanobrassica, and synthetic Brassica napus.
11. Protoplast Fusion
- When sexual hybridization is completely impossible due to pre-fertilisation barriers, protoplast fusion (somatic hybridization) can be used to produce somatic hybrids.
- This is dealt with comprehensively in Part 3. It is listed here as a technique within the context of overcoming wide-cross barriers.
12. Grafting
- Grafting of an interspecific hybrid onto a cultivated species as rootstock has sometimes assisted in making wide crosses successful.
- The grafting relationship may transfer substances that facilitate growth and survival of the otherwise-weak hybrid. This approach is mainly used in ornamentals and some tree species.
1.6 Sterility in Distant Hybrids
Distant hybrids show variable sterility ranging from complete fertility to complete sterility.
- Completely fertile: Lycopersicon esculentum × L. pimpinellifolium hybrid is completely fertile.
- Completely sterile: Sugarcane × maize hybrid is completely sterile.
Distant hybrids can be grouped into two categories:
- Hybrids showing at least some fertility, which can be maintained by selfing, intercrossing, or backcrossing; and
- Hybrids that are completely sterile and must be maintained clonally or by chromosome doubling.
Cytogenetic Basis of Sterility
- Most interspecific hybrids show reduced chromosome pairing at meiosis.
- In extreme cases, all chromosomes may be present as univalents — finding no homologue with which to pair.
- The distribution of univalents at meiosis I is irregular, leading to the formation of unbalanced gametes and partial to complete sterility.
- Additionally, interspecific crosses often show:
- rings and chains at metaphase I (indicating translocations between parental genomes);
- bridges and fragments at anaphase I (indicating inversions);
- loops at pachytene (indicating duplications or deletions).
- All these cytological aberrations reduce fertility. Fertility in such hybrids is improved by doubling the chromosome number to produce amphidiploids.
1.7 Applications of Wide Hybridization in Crop Improvement
1. Alien Addition Lines
- An alien addition line carries the complete chromosome complement of the cultivated species plus one or more additional chromosome pairs from a related wild species.
- The added chromosome(s) carry the desired gene(s).
- These are used for:
- Studying the phenotypic effects of individual alien chromosomes;
- As a step towards gene introgression;
- Direct use where the added chromosome confers the desired trait without major yield penalty.
- Example: Disease resistance genes in wheat, oats, and tobacco have been studied and used through alien addition lines from wild Aegilops, Hordeum, and Nicotiana species.
2. Alien Substitution Lines
- An alien substitution line has one chromosome pair from a wild species replacing (substituting) the corresponding chromosome pair of the cultivated species.
- The substituted chromosome carries the desired gene in a more stable genetic background than an addition line.
- Monosomic substitution analysis (using Sears' wheat nullisomic-tetrasomic series) was the principal method for developing such lines.
3. Introgression of Genes (Chromosome Segment Transfer)
- The most agronomically useful outcome of wide hybridization — transfer of a small chromosome segment carrying the desired gene(s) from a wild species into the cultivated species background through repeated backcrossing.
- After introgression, the cultivated genotype is essentially restored except for the small alien segment with the desired gene.
|
Trait |
Cross (Donor → Recipient) |
Notes |
|
Black arm (bacterial blight) resistance |
G. arboreum → G. barbadense (cotton) |
Interspecific cross within Gossypium genus |
|
Cold tolerance |
Wild relatives → wheat, onion, potato, tomato, grape |
Multiple independent programmes globally |
|
Grassy stunt virus resistance (Bph1) |
O. nivara (wild) → O. sativa (cultivated rice) |
IRRI programme; basis of IR-36's multi-disease resistance |
|
TMV resistance (N gene) |
N. glutinosa → N. tabacum (tobacco) |
Classic early example; N gene from wild Nicotiana |
|
Leaf rust resistance (Lr9) |
Aegilops umbellulata → Triticum aestivum |
Wheat leaf rust resistance from wild Aegilops |
|
Oil quality |
Wild relatives → oil palm |
Source of high-oleic variants |
|
Bhendi (Okra) YVMV resistance |
Abelmoschus manihot → A. esculentus |
Parbhani Kranti variety; see achievements below |
4. Development of New Crop Species
- The most dramatic application — creation of entirely new species through intergeneric hybridization followed by chromosome doubling.
- The new species combines traits of both parents and can be grown as a crop in its own right.
Triticale (× Triticosecale):
- Cross between wheat (Triticum sp.) and rye (Secale cereale).
- First produced by Rimpau in 1890.
- Modern development occurred at CIMMYT (I believe this is approximately correct — please verify institutional details from CIMMYT's published history).
- Hexaploid Triticale (AABBDDRR, 2n=56) combines wheat's grain quality and yield with rye's hardiness, cold tolerance, and resistance to fungal diseases.
- Limitations include poor grain filling and wrinkled grain — substantially improved in modern varieties.
- Grown today in approximately 4 million hectares globally, primarily as animal feed and in some human food uses (verify current area from FAO FAOSTAT).
Brassica napus (canola/oilseed rape):
- An allotetraploid (AACC, 2n=38) derived from crossing B. rapa (AA, 2n=20) and B. oleracea (CC, 2n=18), with subsequent chromosome doubling.
- Now one of the world's most important oilseed crops (canola). This species formed both naturally and through deliberate breeding. The relationship is described as the Triangle of U.
Raphanobrassica:
- Karpechenko (1928) — cross between radish (Raphanus sativus, RR, 2n=18) and cabbage (Brassica oleracea, BB, 2n=18).
- The F1 was sterile (n=9R+9B). After chromosome doubling, a fertile amphidiploid Raphanobrassica (RRBB, 2n=36) was obtained.
- This was the first experimentally produced allopolyploid demonstrating that amphidiploidy could overcome hybrid sterility.
- Agronomically useless — it had the root of a cabbage and the leaves of a radish — but extremely important as a scientific proof of concept.
5. Utilization as New F1 Hybrid Varieties
In crops where vegetative propagation is possible, or where seed setting in F1 hybrids is adequate, interspecific F1 hybrids can be directly used as commercial varieties:
- Cotton: Varalaxmi cotton variety — an F1 hybrid derived from Gossypium hirsutum × G. barbadense — combines the adaptability of G. hirsutum with the long staple fibre quality of G. barbadense. (Verify current status and exact parentage from CICR, Nagpur.)
- Sugarcane: All present-day commercial sugarcane varieties are complex interspecific hybrids primarily involving Saccharum officinarum (noble cane — high sugar, low fibre) and S. spontaneum (wild cane — hardy, disease-resistant, ratoons well). The process of nobilization (crossing S. officinarum with S. barberi or S. spontaneum followed by backcrossing to recover high sugar) was pioneered by T.S. Venkatraman and C.A. Barber in India.
1.8 Achievements of Wide Hybridization in India
|
Variety |
Cross |
Achievement |
|
Parbhani Kranti (Bhendi/Okra) |
Abelmoschus esculentus cv. Pusa Sawani × A. manihot |
Resistant to yellow vein mosaic virus (YVMV); Kharif yield approximately 110-120 q/ha; Summer approximately 85-90 q/ha. Note: verify exact yield figures from ICAR-IIVR or Maharashtra sources. |
|
Pusa Kranti (Bhendi) |
Similar A. esculentus × A. manihot cross |
Kharif yield approximately 105-110 q/ha; Summer approximately 75-80 q/ha. Same verification note applies. |
|
Sugarcane Co varieties |
S. officinarum × S. spontaneum and S. barberi |
Nobilization programme at SBI Coimbatore; all commercial Indian sugarcane varieties descend from these crosses |
|
Varalaxmi Cotton |
G. hirsutum × G. barbadense |
Long staple hybrid; verify current cultivation status |
|
Wheat rust-resistant lines |
T. aestivum × Aegilops species |
Multiple leaf rust (Lr) and stem rust (Sr) resistance genes introgressed; foundation of rust-resistant Indian wheats |
|
Rice CO 31 |
O. sativa var. indica × O. perennis |
An early interspecific cross; verify details from TNAU rice records |
1.9 Limitations of Distant Hybridization
- Incompatible crosses: Many wide crosses fail at various stages from pollen germination to embryo development.
- F1 sterility: Even when hybridization succeeds, the F1 is often completely or partially sterile due to chromosome pairing failure.
- Problems in creating new species: Most newly created allopolyploids have defects — low fertility, cytogenetic instability, undesirable traits from the wild parent — and require extensive further breeding to become agronomically useful.
- Lack of homeology: When the chromosomes of the two parental species have no structural homology, chromosome recombination is impossible and the introgression of specific gene segments becomes very difficult.
- Undesirable linkages (linkage drag): Genes responsible for undesirable wild-type characters (late maturity, shattering, low yield, poor quality) may be tightly linked to the desired resistance gene on the alien chromosome segment. Breaking such linkages requires many backcross generations and large population sizes.
- Difficulty with recessive and quantitative traits: Transfer of recessive genes requires additional selfing generations; transfer of quantitative (polygenic) traits from wild relatives is even more difficult because the individual gene contributions are small and may be disrupted by the cultivated parent background.
- Lack of flowering in F1: Some interspecific F1 hybrids fail to flower.
- Improved varieties as parents: As cultivated varieties become more genetically improved, the genetic gap between cultivated varieties and wild relatives increases, making wide crosses progressively more difficult.
- Dormancy: Wild relatives often have strong seed dormancy. Seeds may not germinate readily in the breeding nursery, making it difficult to raise the wild parent for hybridization.
Key Examples of Distant Hybridization
|
New Variety / Product |
Female Parent |
Male Parent |
Key Achievement |
|
Triticale |
Triticum sp. |
Secale cereale (rye) |
New crop species; wheat quality + rye hardiness |
|
Raphanobrassica |
Raphanus sativus (radish) |
Brassica oleracea (cabbage) |
First man-made allopolyploid; no agricultural value |
|
Brassica napus |
B. rapa (AA) |
B. oleracea (CC) |
Major oilseed crop; canola originated from this cross |
|
Parbhani Kranti (bhendi) |
A. esculentus |
A. manihot |
Yellow vein mosaic resistance in okra |
|
CO 31 Rice |
O. sativa var. indica |
O. perennis |
Early interspecific rice variety |
|
Swarna-Sub1 |
Swarna (O. sativa) |
FR13A (O. sativa) |
Submergence tolerance via Sub1A gene — same species technically |
|
Tobacco TMV resistance |
N. tabacum |
N. glutinosa |
N gene for TMV resistance introgressed into tobacco |
|
Wheat leaf rust (Lr9) |
T. aestivum |
Aegilops umbellulata |
Lr9 gene for leaf rust resistance from wild Aegilops |