PB Ch 35. Hybridization
1.1 Definition of Hybridization
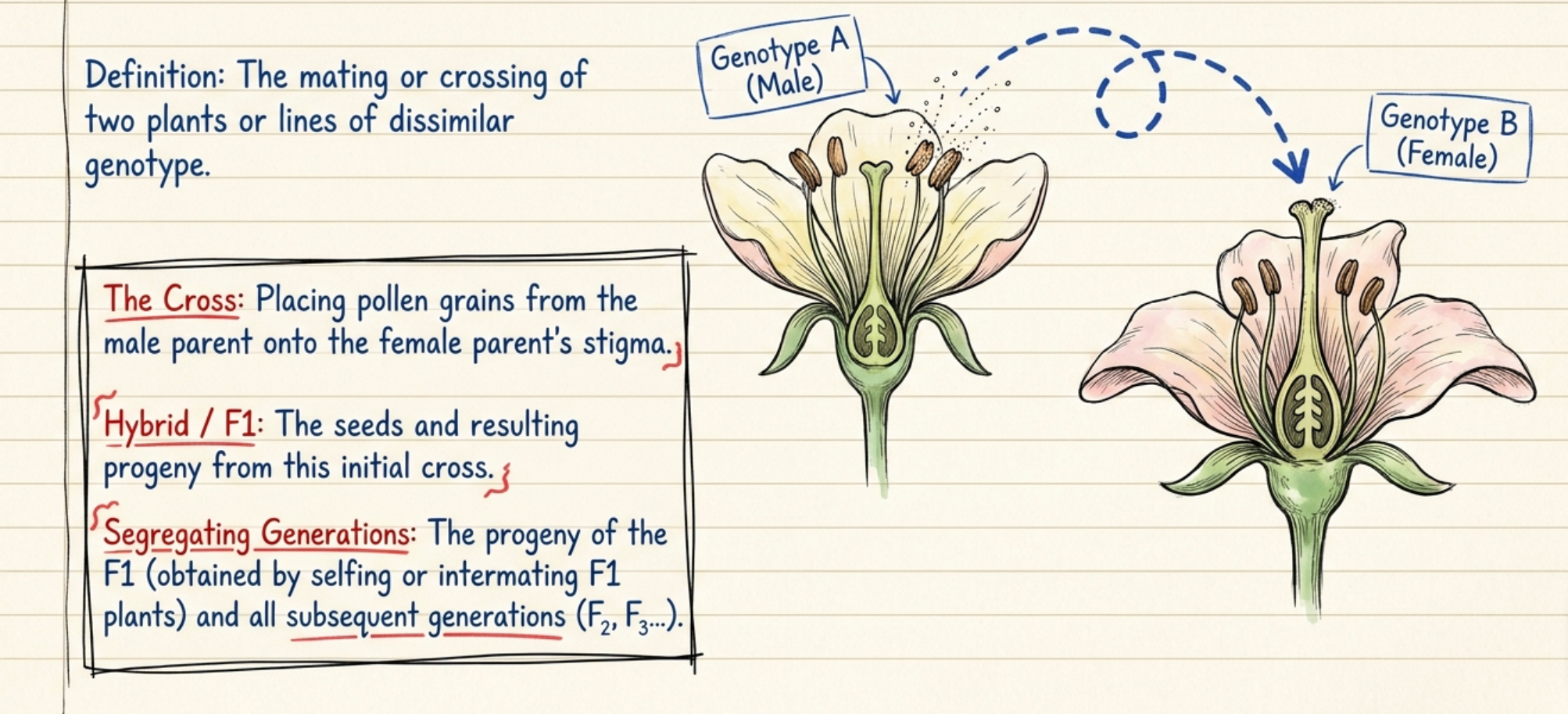
- The mating or crossing of two plants or lines of dissimilar genotype is known as hybridization.
- In plants, crossing is performed by placing pollen grains from one genotype — the male parent — on to the stigma of flowers of the other genotype — the female parent.
- The seeds as well as the progeny resulting from hybridization are known as the hybrid or F1.
- The progeny of the F1, obtained by selfing or by intermating F1 plants, and the subsequent generations are termed segregating generations.
- The first natural hybridization was recorded by Cotton Mather (1716) in corn.
- The first artificial interspecific plant hybrid was produced by Thomas Fairchild in 1717, commonly called Fairchild's Mule.
- Hybridization was first practically used in crop improvement by the German botanist Joseph Koelreuter in 1760.
- After Mendel, hybridization became the central method of crop improvement, and the vast majority of modern crop varieties have resulted from hybridization.
1.2 Objectives of Hybridization
- The chief objective of hybridization is to create genetic variation.
- When two genotypically different plants are crossed, genes from both parents are brought together in F1.
- Segregation and recombination produce many new gene combinations in F2 and later generations. Three major objectives are:
A. Combination Breeding
- The main aim here is to transfer one or more characters into a single variety from other varieties. These characters may be governed by oligogenes or polygenes.
- In combination breeding, genetic divergence between parents is not the major consideration — what matters is that one parent must possess the character(s) under transfer in sufficient intensity, while the other is generally a popular, well-adapted variety.
- The backcross method of breeding was designed for combination breeding; the pedigree method also frequently fulfils this purpose.
- Example: Transfer of rust resistance from a donor line into the popular variety HD 2967 using the backcross method — the donor provides the resistance gene(s), while HD 2967 is the recurrent parent providing all other agronomic background.
B. Transgressive Breeding
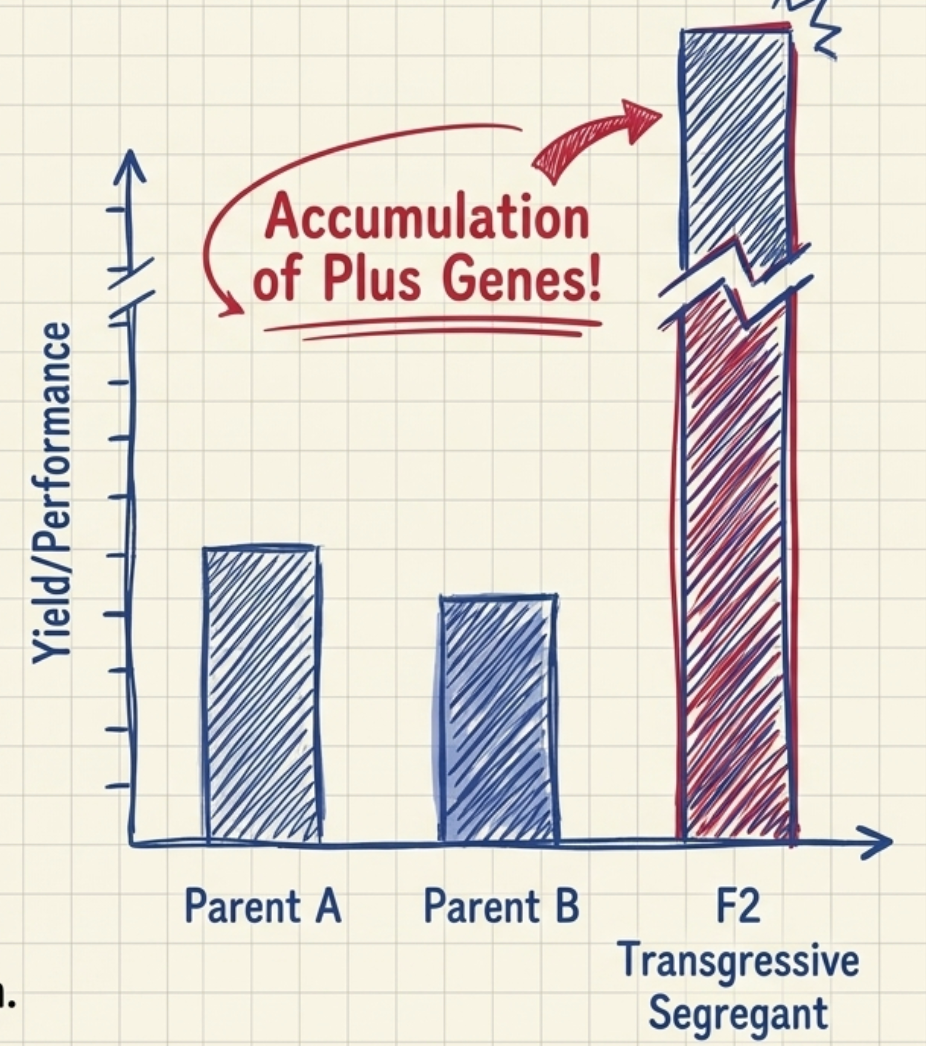
- Transgressive breeding aims to improve yield or its component characters through transgressive segregation.
- Transgressive segregation is the production of plants in the F2 generation that are superior to both parents for one or more characters.
- Such plants are produced by accumulation of plus (favourable) genes from both parents — genes that recombine to give a segregant exceeding either parent.
- The pedigree method and the population approach are especially designed for recovering transgressive segregants.
- For transgressive segregation to occur, both parents must possess different sets of plus genes; parents must combine well and should preferably be genetically diverse. This is why parent selection and evaluation of parental diversity (using DUS analysis or molecular markers) is critical.
C. Production of Hybrid Varieties
- In most self-pollinated crops, the F1 is more vigorous and higher-yielding than the parents. Where it is commercially feasible, F1 may be used directly as a variety.
- In such cases, the two parents must produce an outstanding F1 — this is assessed through combining ability analysis (test crosses and topcrosses to estimate GCA and SCA).
- Hybrid variety development requires maintaining the two parental inbred lines and producing F1 seed each season, which is the central challenge of hybrid seed production.
1.3 Types of Hybridization
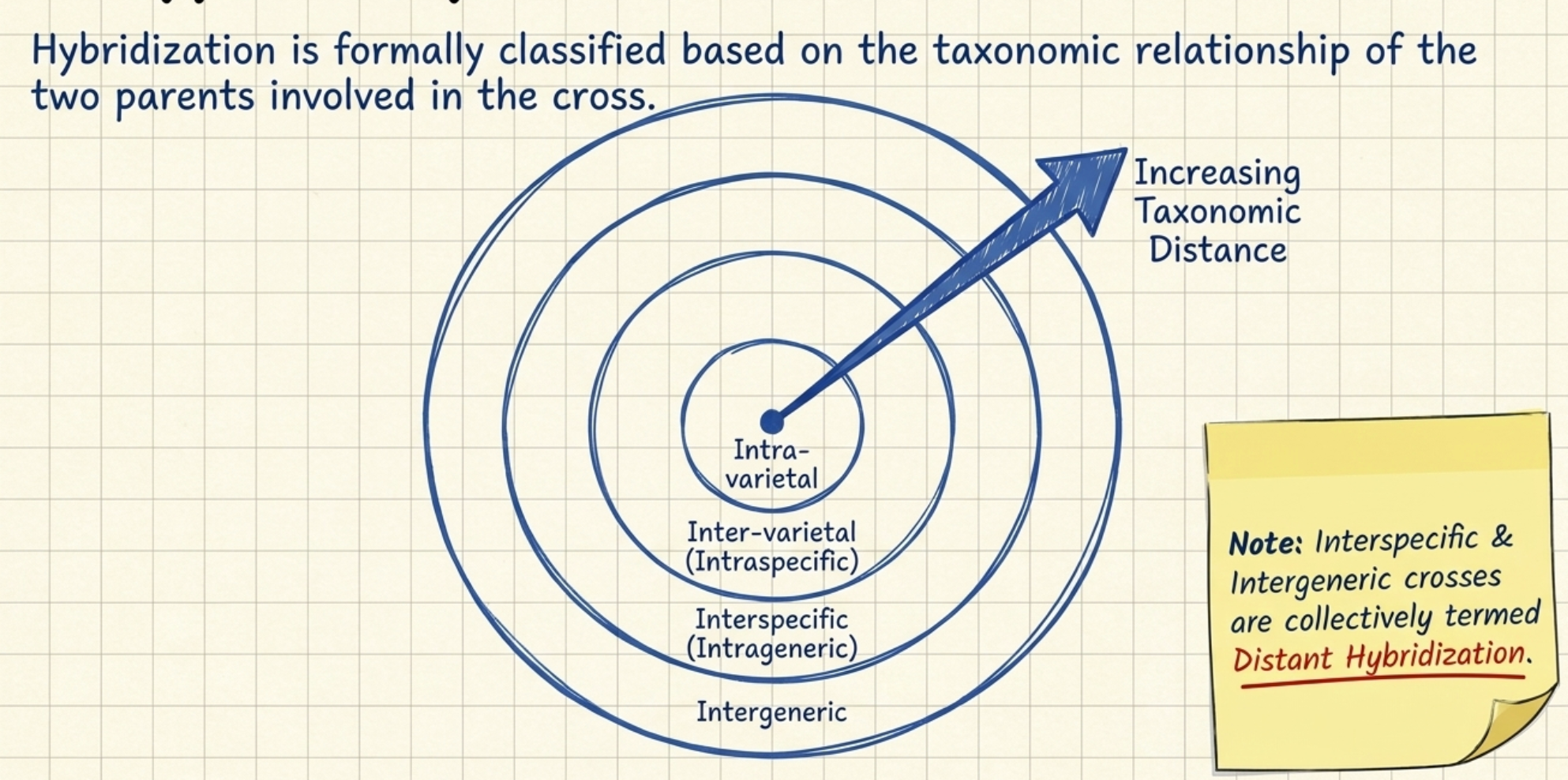
Based on the taxonomic relationship of the two parents involved, hybridization is classified as follows:
(i) Intra-varietal Hybridization
- Crosses are made between plants of the same variety. Both parents belong to the same variety but differ in one or a few characters due to natural or induced variation within the variety.
- This is used to combine minor variation within a variety.
(ii) Inter-varietal (Intraspecific) Hybridization
- Crosses are made between plants belonging to two different varieties of the same species.
- This is the most commonly used form of hybridization in crop improvement worldwide.
- It is so common that it often appears to be the only form used.
- Examples: crossing two varieties of wheat, or two varieties of rice.
- Inter-varietal crosses may be simple or complex depending on the number of parents involved:
- Simple Cross: Two parents crossed to produce F1. A × B → F1(A×B). F1 is then selfed to produce F2 or used in a backcross programme.
- Complex (Convergent) Cross: More than two parents crossed to bring together genes from several parents into one hybrid. Example of a four-parent complex cross: (A × B) × (C × D) → Complex hybrid. These crosses are now routine in advanced self-pollinated crops like wheat and rice.
(iii) Interspecific Hybridization (Intrageneric)
- Crosses are made between two different species of the same genus.
- Example: Oryza sativa × O. perennis (for developing CO 31 rice); Gossypium hirsutum × G. barbadense (for long-staple cotton improvement).
(iv) Intergeneric Hybridization
- Crosses are made between species belonging to different genera.
- Example: Triticum sp. × Secale cereale (rye) → Triticale.
- These crosses are collectively termed distant hybridization and are dealt with comprehensively in Part 2 of this chapter.
(v) Introgressive Hybridization
- Transfer of some genes from one species into the genome of another species is known as introgressive hybridization.
- It is essentially repeated backcrossing after an initial wide cross, resulting in a cultivated variety that has received a small chromosomal segment from a wild relative carrying the desired gene.
- The rest of the genome reverts to the recurrent (cultivated) parent background. Examples include transfer of Lr9 (leaf rust resistance) from Aegilops umbellulata into wheat.
Interspecific vs Intergeneric Hybridization
|
Feature |
Interspecific Hybridization |
Intergeneric Hybridization |
|
Parents belong to |
Same genus, different species |
Different genera |
|
Taxonomic distance |
Closer (within one genus) |
Greater (between genera) |
|
Crossability |
Moderate to good |
Very low; major barriers |
|
F1 sterility |
Partial to complete |
Usually complete |
|
Example |
O. sativa × O. nivara (rice) |
Triticum × Secale (Triticale) |
|
Embryo rescue needed |
Sometimes |
Almost always |
|
New species possible |
Possible (amphidiploids) |
Yes (e.g., Triticale, B. napus) |
1.4 Procedure of Hybridization — Steps
The standard procedure for hybridization involves the following sequential steps. The details of each step depend on the crop's floral biology.
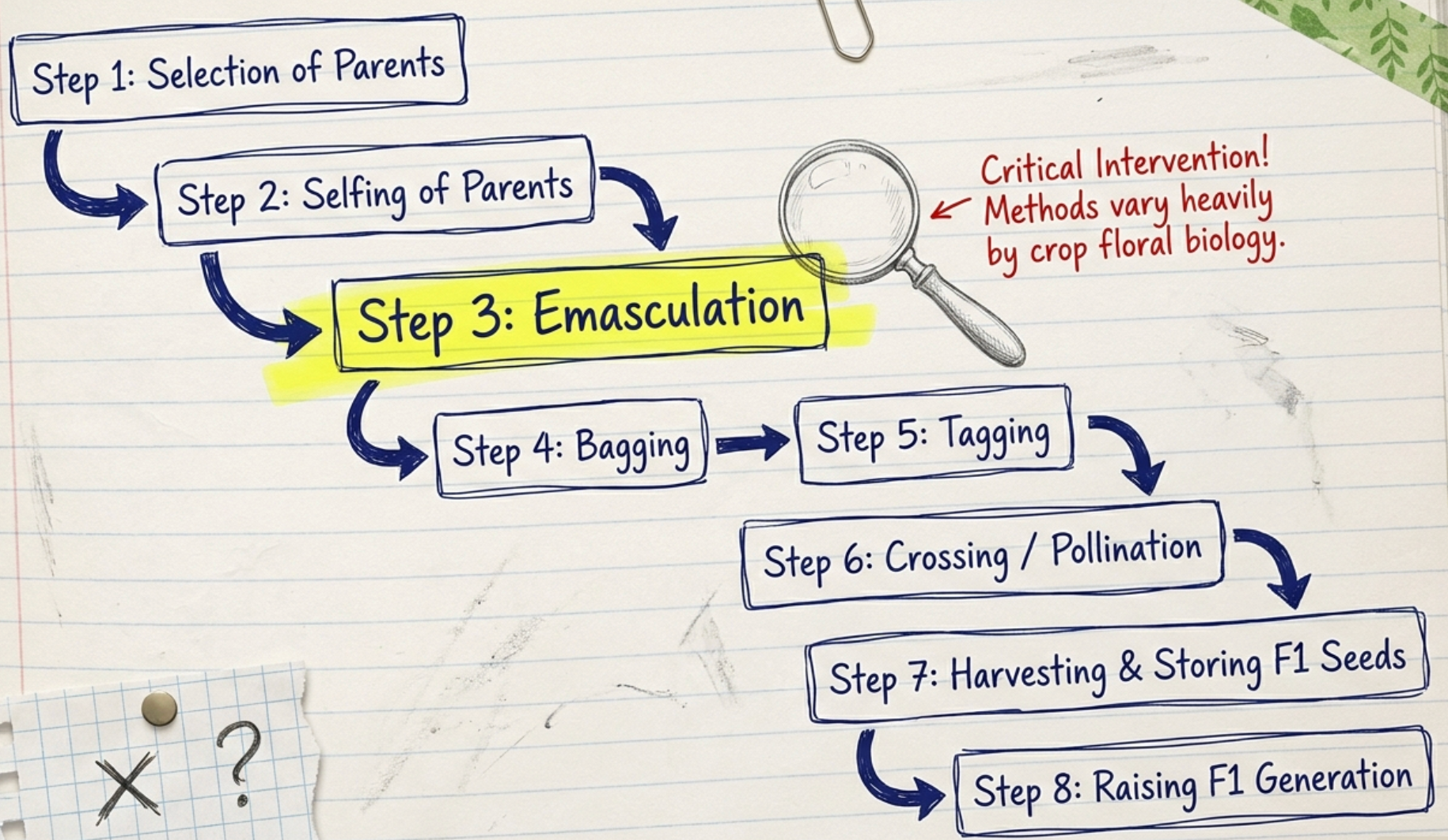
Step 1: Selection of Parents
Parents must be selected on the basis of the objectives of the breeding programme. Key criteria:
- Complementarity: Parents should complement each other — each should have characters the other lacks.
- Genetic diversity: For transgressive breeding, parents should be genetically diverse. For combination breeding, one parent can be the popular adapted variety.
- Adaptability: Parents should be reasonably well-adapted to the local conditions, especially the recurrent parent in a backcross programme.
- Combining ability: Where feasible, test parents for GCA through topcrosses or diallel analysis before committing to large-scale hybridization.
Step 2: Selfing of Parents
- Parents are selfed for one to two seasons before crossing.
- This induces homozygosity, eliminates undesirable recessive characters, and ensures that each parent breeds true.
- In cross-pollinated species, parents are developed as inbred lines through repeated self-pollination.
Step 3: Emasculation
- Emasculation is the removal of stamens or anthers, or the killing of pollen, from the flower of the female parent before they mature and shed pollen — without damaging the female reproductive organs in any way.
- This prevents self-pollination on the female parent.
- Emasculation is not required in:
- Unisexual plants — where staminate and pistillate flowers are separate (monoecious: maize, castor; dioecious: papaya, date palm).
- Plants where male sterility (GMS, CMS, or CGMS) is used — the male-sterile line serves as the female parent and no manual emasculation is needed.
- Plants showing effective self-incompatibility — pollen from same plant does not fertilise own stigma.
Where emasculation IS required (bisexual, self-pollinated plants), the following methods are used:
Methods of Emasculation
(a) Hand Emasculation / Forceps Method: Used in plants with large flowers — cotton, pulses, wheat. The corolla is gently opened and anthers are carefully removed using fine-tip forceps before they dehisce. Done generally between 4–6 PM the day before anthers are expected to shed pollen. Stigma and ovary must not be damaged.
(b) Hot Water Method: Used in small-flowered crops — sorghum, pearl millet, rice. The flower/panicle is immersed in hot water for a defined period at a temperature that kills pollen but does not damage the gynoecium.
|
Crop |
Temperature |
Duration |
Notes |
|
Rice |
40–44°C |
5–10 minutes |
Thermos flask used; Rhind's or Dr. Ramaiah method |
|
Sorghum |
48°C |
10 minutes |
Plastic bag / mass emasculation technique also used |
|
Pearl millet |
Not required |
— |
Protogyny allows crossing without emasculation |
|
Ragi (Finger millet) |
52°C |
2 minutes |
Hot water treatment best for small florets |
(c) Cold Water Method: In rice, 0–6°C temperature kills pollen grains without damaging gynoecium. Less effective than hot water treatment. Used as an alternative where hot water method is not feasible.
(d) Alcohol Treatment: In lucerne (alfalfa), 57% alcohol for 10 seconds kills pollen. Duration must be precise — longer treatment can damage the gynoecium. Rarely used now due to difficulty in standardising exposure time.
(e) Suction / Vacuum Emasculation Method: Used in crops with minute flowers. Suction pressure removes anthers while leaving the pistil intact. In vacuum emasculation (developed at CRRI, Cuttack for rice), six panicles can be emasculated simultaneously. Hand emasculation yields about 100 flowers per person per hour; vacuum emasculator allows 3000–3600 florets per hour when six persons operate. Note: approximately 10–15% selfing may still occur.
(f) Chemical Gametocides: Certain chemicals induce male sterility when sprayed before flowering. Examples: Maleic hydrazide (MH), 2,4-D, NAA (naphthalene acetic acid), tribenzoic acid, ethrel (ethephon), FW-450 (used in cotton), RH-531. The optimal dose and timing must be precisely calibrated — too high or too early can damage the crop; too late is ineffective.
(g) Genetic Male Sterility or Self-Incompatibility: Plants carrying male-sterile genes or self-incompatibility alleles serve as female parents without any emasculation. Widely used in commercial hybrid seed production in sorghum, pearl millet, sunflower, rice (CGMS system), and vegetables.
Crop-Specific Emasculation and Crossing Techniques
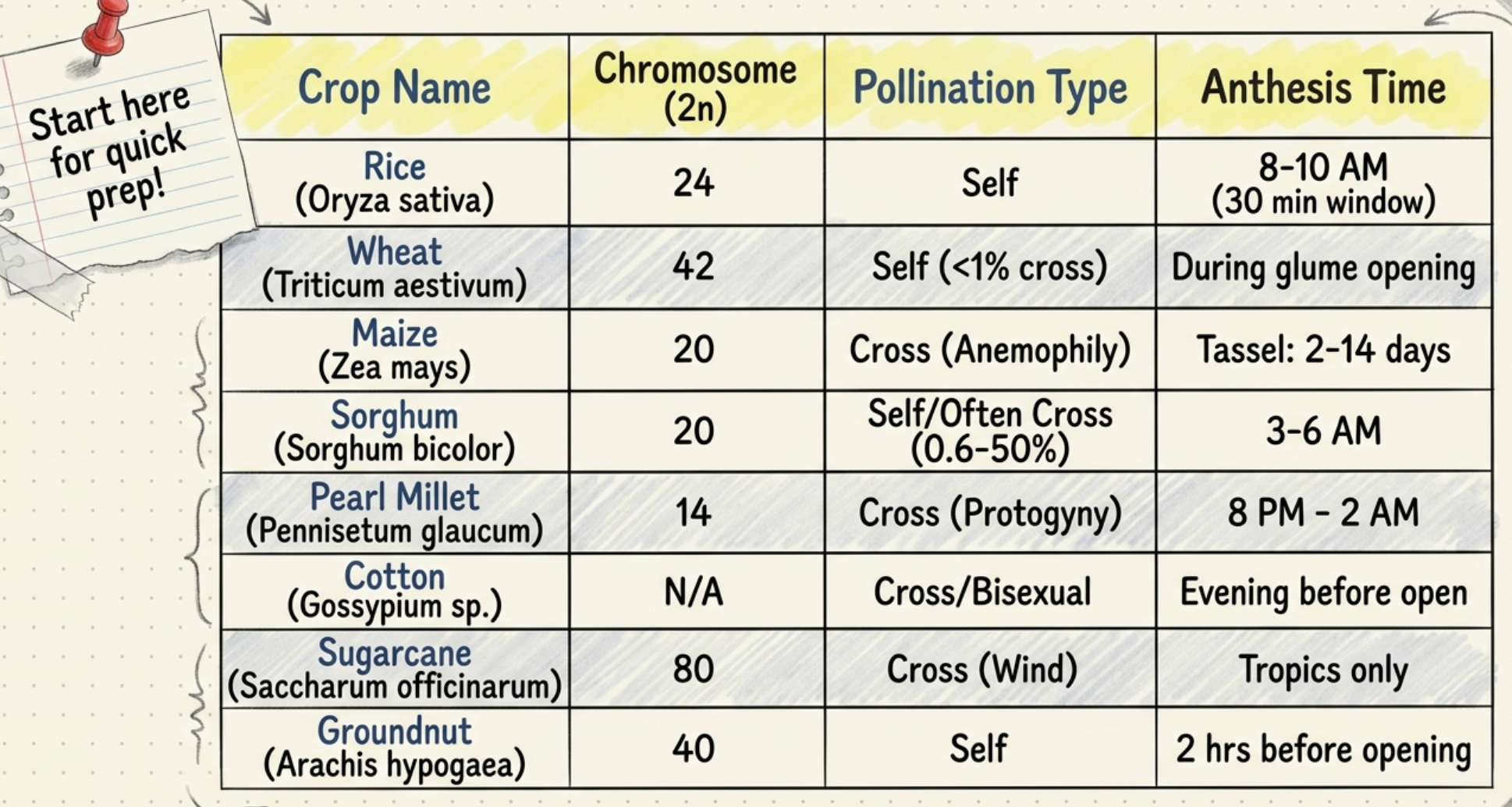
RICE (Oryza sativa, 2n = 24):
- Anthesis commences shortly after panicle emergence. Spikelets at the tip bloom first and proceed downwards. Anthesis time: 8–10 AM. Each spikelet remains open for about 30 minutes. Anther dehiscence takes place immediately after the spikelets open. Stigma remains receptive for one day.
- Clipping method: In the previous evening, top 1/3 and bottom 1/3 of the panicle are clipped off with scissors, leaving middle spikelets. The top 1/3 of each remaining spikelet is clipped in a slanting position. The six anthers are then removed with a needle. The panicle is covered with a butter paper bag. Next morning (9 AM), the male parent panicle is inserted inverted into the bag and turned to disperse pollen.
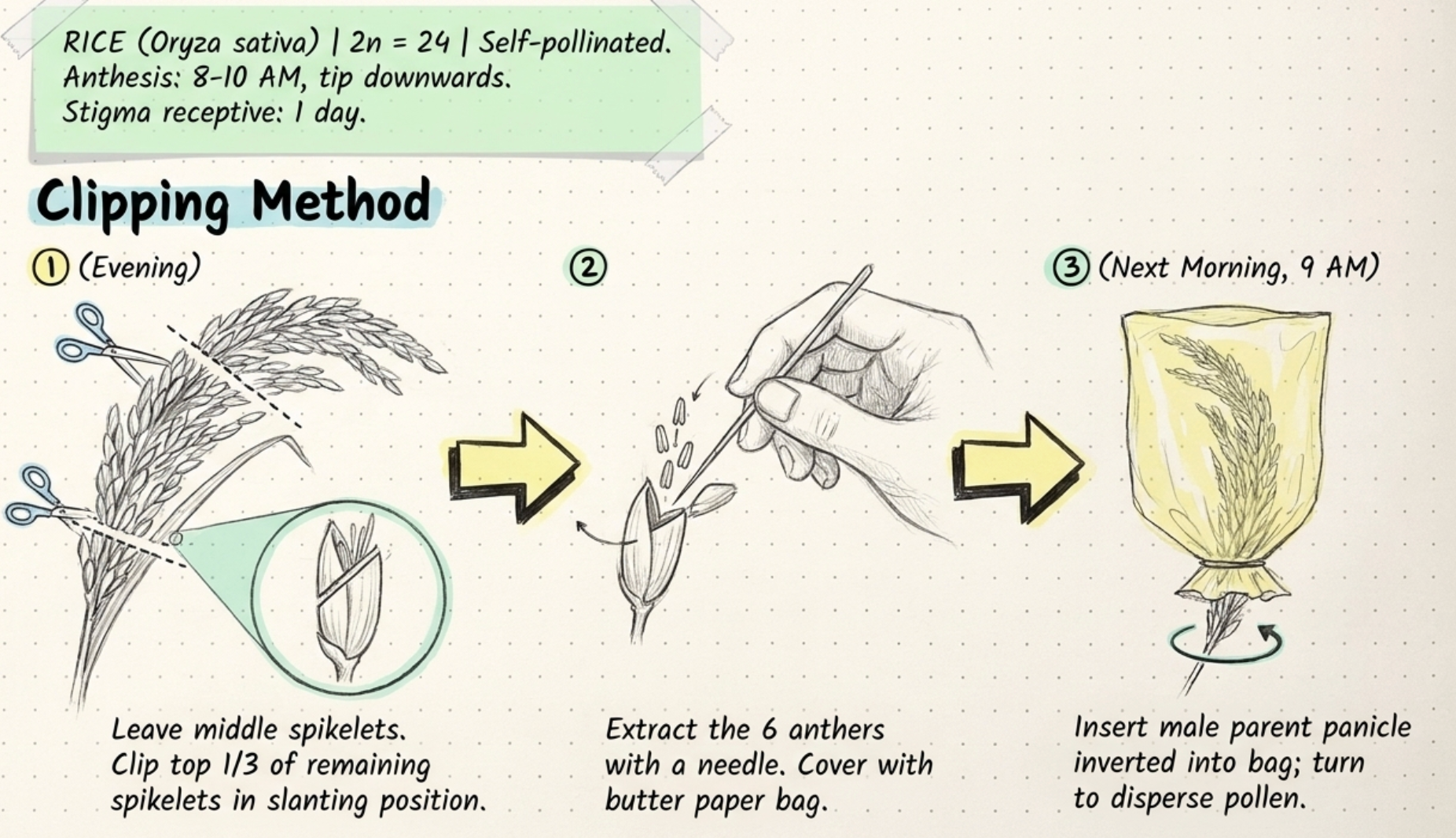
- Hot water method: Panicles on 3rd or 4th day of blooming selected. One hour before blooming (around 7 AM), undeveloped and already-opened spikelets are removed. The panicle is immersed in a thermos flask at 40–44°C for 5–10 minutes. Then the six stamens are removed with forceps.
- Dr. Ramaiah method: Panicles on 3rd or 4th day selected. Top and lower spikelets removed. Panicle covered with wet cloth and air blown through the mouth — this causes the spikelets to open within 2–3 minutes. Six anthers then removed.
- Vacuum emasculation method: Works on the principle of suction pressure. Spikelets are clipped; the minute pipette is applied at the clipping point and pollen is sucked in. Developed at CRRI, Cuttack. Rate: approximately 3000–3600 florets per hour with six operators vs 100 by hand. Note: verify exact rate from CRRI publications.
- Cuttack method: Panicle inserted into hollow bamboo closed at one end. Flowers open within 5–10 minutes inside the bamboo. Anthers then removed.
- Brown paper method: Panicles enclosed in brown paper cover a couple of hours before blooming. Heat inside causes anther extrusion without dehiscence. After 15–30 minutes anthers are clipped off and pollen from male parent dusted on stigma.
- Rhind's method: Hot water flask inverted; residual warmth and humidity inside the flask causes spikelets to open. Anthers exposed and removed with forceps.
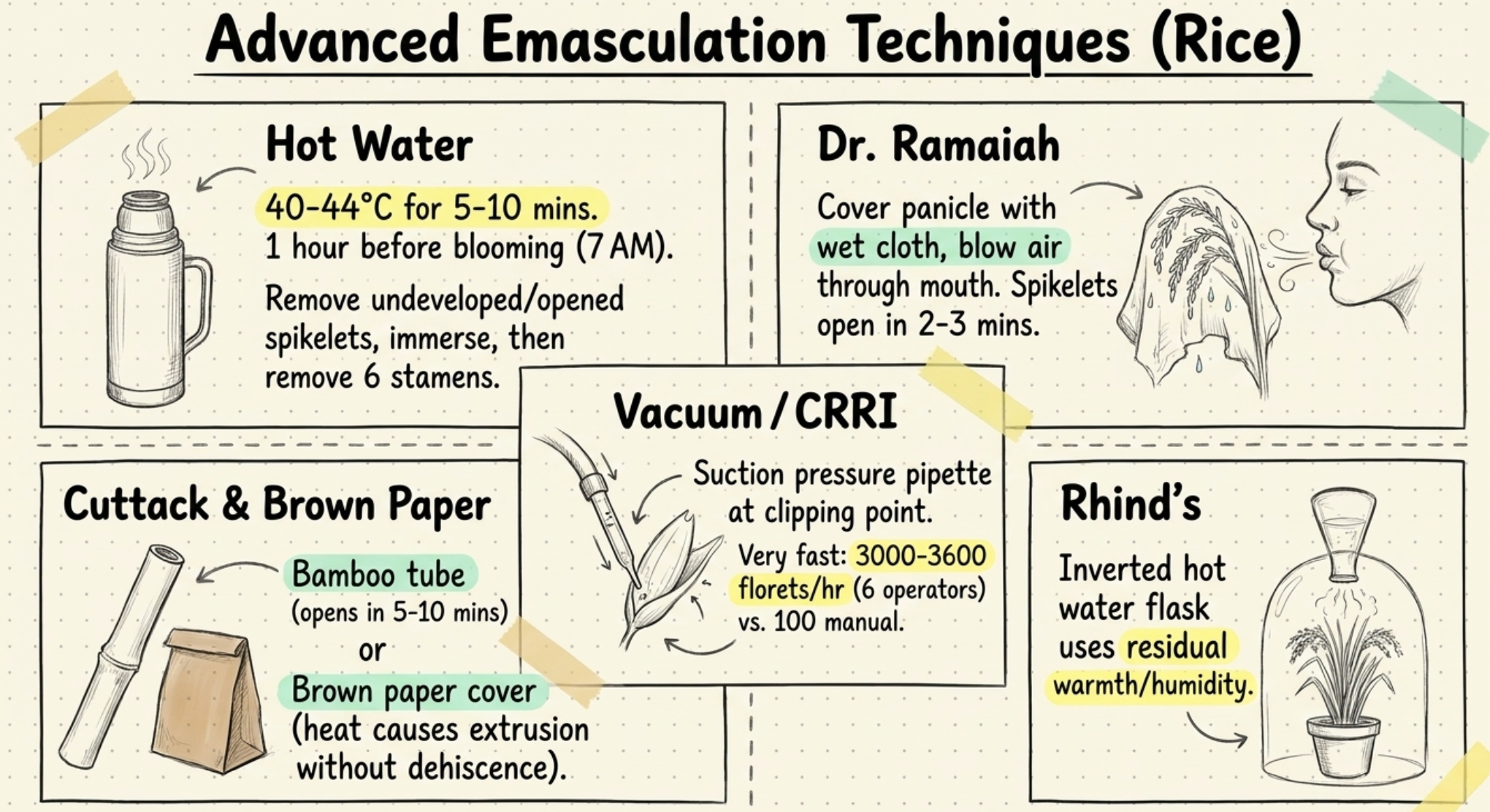
WHEAT (Triticum aestivum, 2n = 42):
- Wheat is largely self-pollinated.
- The glumes open during flowering; anthers protrude from glumes and part of the pollen is shed outside, but entry of foreign pollen results in only minor cross-pollination normally less than 1 per cent.
- Emasculation is done by: removing upper 1/3 of the spikelet, removing lower spikelets and alternate spikelets on both sides.
- The remaining anthers are carefully exposed and removed. The emasculated ear is covered with butter paper. Next day, ripe bright yellow anthers from the male parent are rubbed on the styles.
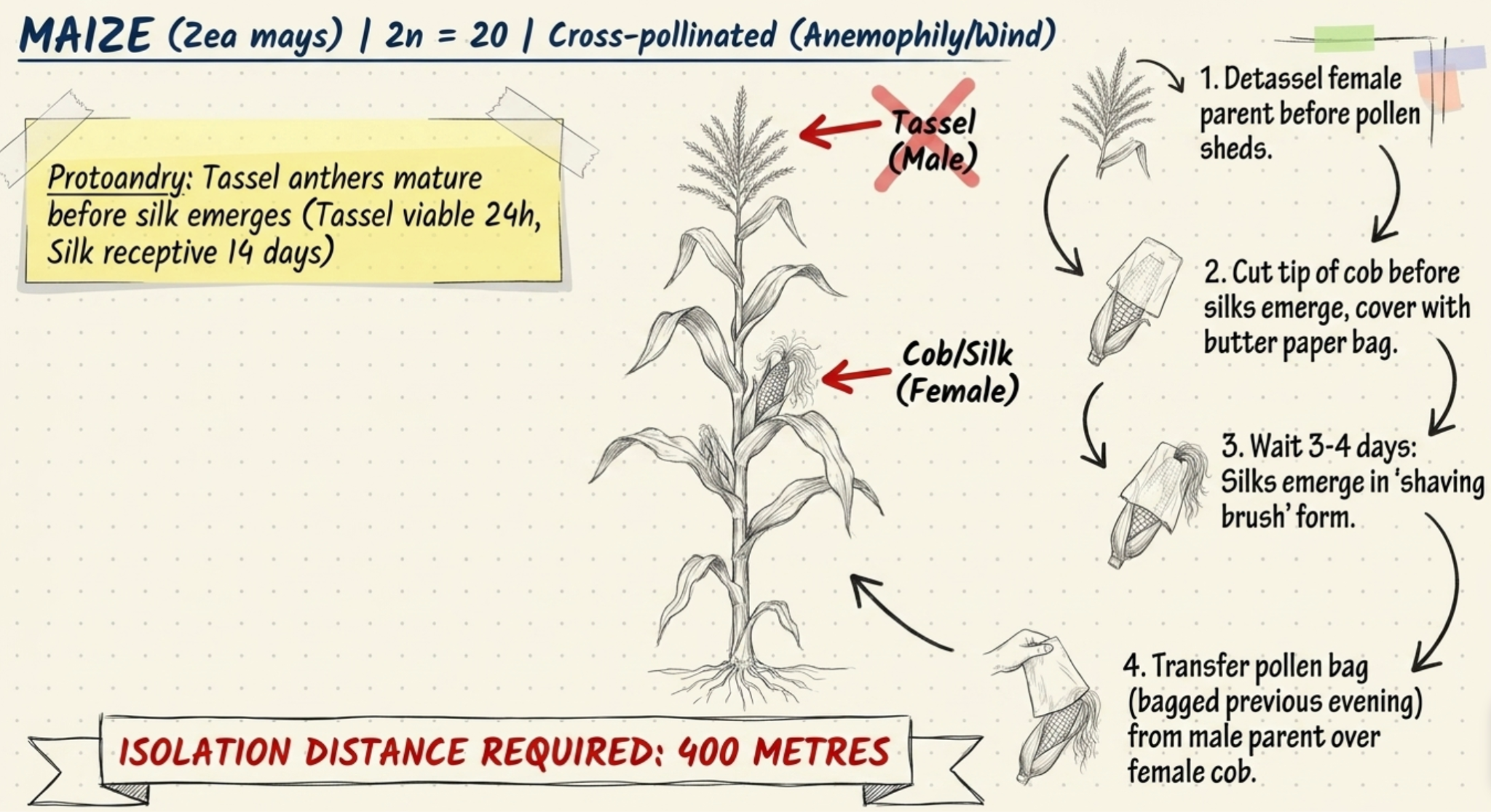
MAIZE (Zea mays, 2n = 20):
- Maize is predominantly cross-pollinated.
- Wind pollination (anemophily) is the general rule.
- It has a monoecious inflorescence with separate male (tassel) and female (cob) flowers.
- Protoandry means the tassel anthers mature before the silk emerges, ensuring natural cross-pollination.
- Tassel anthesis extends 2–14 days; pollen viability remains for about 24 hours. The stigma (silk) is receptive throughout its length for up to 14 days.
- Crossing technique:
- The tassel of the female parent is removed (detasseling) before it sheds pollen.
- The tip of the cob is cut before silks emerge and covered with a butter paper bag. After 3–4 days, silks emerge in a 'shaving brush' form. The male parent tassel is bagged the previous evening.
- Pollen is transferred by placing the male parent pollen-containing bag over the female cob after removing the cob cover. The isolation distance for maize is 400 metres.
SORGHUM (Sorghum bicolor, 2n = 20):
- Sorghum is normally self-pollinated but some florets are protogynous, resulting in cross-pollination averaging about 6 per cent.
- The amount of natural cross-pollination varies from 0.6 to 50 per cent depending on variety and location — more in loose panicles than compact ones.
- Anthesis starts from tip downwards at 2–5 cm per day and completes within 7–10 days. Anthesis time: 3–6 AM. Pollen is viable for a short period; stigma is receptive for 8–16 hours.
- Three emasculation methods are used :
- Hand emasculation — needle inserted at middle of floret, moved across glumes and rotated 90° to lift out anthers; about 50 florets emasculated per set.
- Hot water method — 48°C for 10 minutes on clusters of 2–3 florets.
- Plastic bag / mass emasculation technique — sorghum panicle enclosed in plastic bag creating high humidity; florets open and anthers emerge but shed no pollen; anthers knocked free by tapping.

PEARL MILLET / CUMBU / BAJRA (Pennisetum glaucum, 2n = 14):
- Naturally cross-pollinated. Protogyny is the key mechanism: stigmas emerge first, then anthers from perfect florets, followed by anthers from staminate florets.
- Anthesis time: 8 PM to 2 AM. Pearl millet does NOT require emasculation for controlled crosses because of protogyny — the female line is covered before stigma emergence.
- After most stigmas have emerged, pollen from the desired male is dusted.
- Another method: the top of the cover of the male inflorescence is clipped and inserted to brush the stigma. Crossed heads collected after 30–35 days.
COTTON (Gossypium sp.):
- Cotton has large bisexual flowers — hand emasculation is straightforward and accurate. Emasculation is done in the evening before the flower opens.
- Mature buds are easily identified. The standard and wing petals are opened; staminal column carefully removed.
- Stigma dusted with pollen next morning (7–10 AM). In large flowers like cotton, petals may naturally fold down the sexual organs, so manual assistance is needed to ensure pollen reaches the stigma.
- Chemical gametocide FW-450 is also used in cotton to induce male sterility when large-scale hybrid seed production is needed.
SUGARCANE (Saccharum officinarum, 2n = 80):
- Cross-pollination is the rule.
- Self male and female sterility, protogyny, and hanging anthers away from stigma at anthesis promote outcrossing.
- Natural pollination is by wind.
- Hybridization in sugarcane is very difficult for four reasons:
- Many varieties seldom flower outside the tropics;
- Spikelets are minute so hand emasculation is impossible;
- Self-sterility of both pollen and ovule predominates;
- Hot water treatment is not possible. Special methods are therefore used:
• Coimbatore method: Cut stems with 1–2 buds are placed in moist mud; roots develop within 10 days. Male and female plants are brought into a crossing block and covered with a common lantern (bamboo frame covered with muslin cloth or polythene). Free pollen shed from the male falls on the female. Both selfed and crossed seeds are harvested from the female; selfed seeds identified by chromosome count in the nursery.
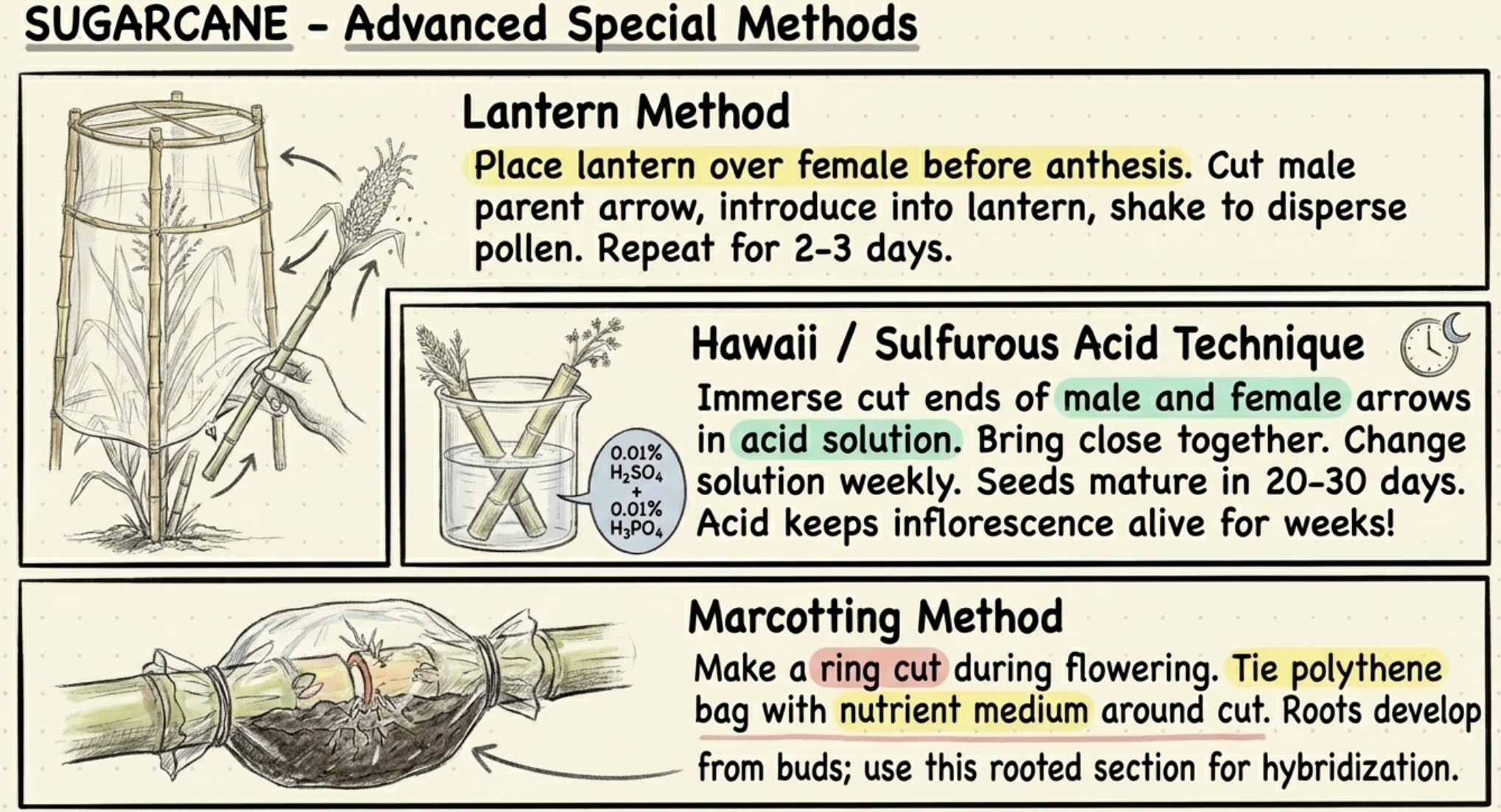
• Lantern method: A lantern is placed over the female plant before anthesis. The male parent's flowering arrow is cut and introduced into the lantern, then shaken to disperse pollen. Repeated for 2–3 days.
• Hawaii / Sulfurous acid technique: Both male and female flowering arrows are cut and their cut ends immersed in 0.01% sulfuric acid + 0.01% phosphoric acid solution. The cut ends are brought close together for cross-pollination. Solution changed weekly; seeds mature in 20–30 days. The acid keeps the inflorescence alive and functional for several weeks.
• Marcotting method: A ring cut is made around the stem during flowering; a polythene bag with nutrient medium is tied around the cut. Roots develop from buds near the cut end. This rooted section is then used for hybridization.

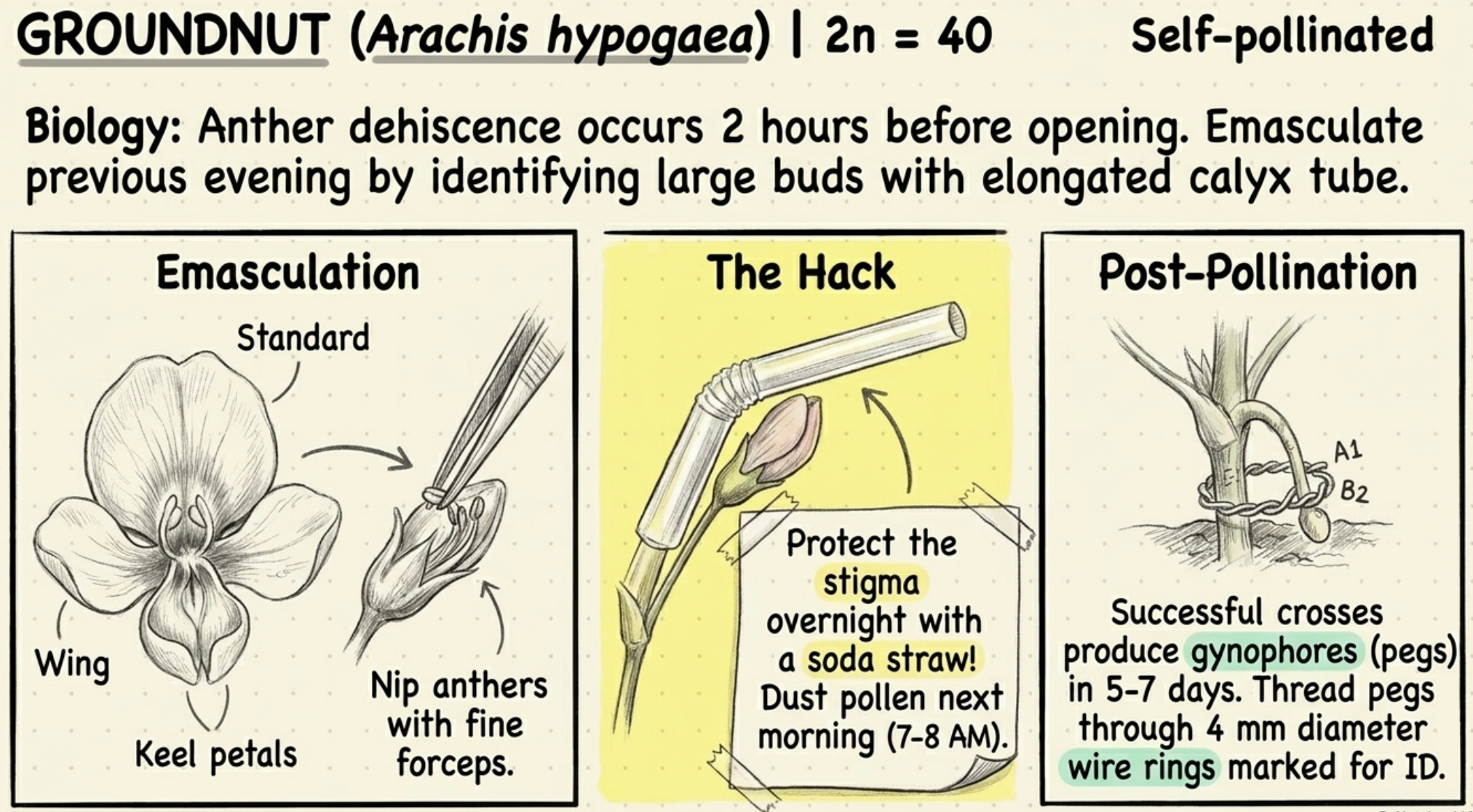
GROUNDNUT (Arachis hypogaea, 2n = 40):
- Self-pollination is the rule.
- Anther dehiscence occurs two hours before the flower opens.
- Emasculation done the previous evening by identifying large buds with elongated calyx tube.
- The standard, wing and keel petals are carefully removed to expose the stamens.
- Anthers nipped off with fine forceps.
- A soda straw is placed over the emasculated bud to protect the stigma.
- Next morning (7–8 AM) pollen is dusted from male parent.
- Successful crosses produce gynophores (pegs) 5–7 days after pollination.
- Pegs are threaded through small wire rings (4 mm diameter) marked for identification.
Step 4: Bagging
- The emasculated flower or inflorescence is immediately covered with a bag to prevent contamination by foreign pollen.
- Bags may be made of butter paper (vegetable parchment), glassine paper, or fine muslin cloth. Butter paper bags are most commonly used as they allow some air circulation while preventing pollen entry.
- Bags are tied at the base of the inflorescence with thread or wire. In unisexual plants, both male and female flowers are bagged separately — female flowers to prevent cross-pollination, male flowers to collect pollen.
Step 5: Tagging
- Emasculated flowers are tagged immediately after bagging. Tags (circular or rectangular, approximately 3 × 2 cm) are attached at the base of the flower or inflorescence.
- The tag must record:
- Field record number;
- Date of emasculation;
- Date of crossing;
- Parentage — female parent written first, then × sign, then male parent (e.g., C × D means C is female, D is male).
Step 6: Crossing / Pollination
- Mature, viable pollen is collected from the male parent and placed on the receptive stigma of the emasculated female flower.
- Pollen is collected in petri dishes (wheat, cotton) or paper bags (maize).
- Application is done using a camel-hair brush, small piece of paper, toothpick, or forceps depending on the crop.
- In sorghum and pearl millet, the male and female inflorescences are enclosed in the same bag to allow free pollen transfer.
- Timing is critical — pollen should be applied at peak stigma receptivity, generally in the morning (8–10 AM for most crops).
Step 7: Harvesting and Storing F1 Seeds
- Crossed pods, heads, or bolls are harvested after maturity and dried thoroughly.
- Seeds are threshed and stored with the original tags intact to maintain cross identity.
- Each crossed family must be kept separate and properly labelled.
Step 8: Raising the F1 Generation
- Stored F1 seeds are sown the following season to raise the F1 generation.
- All plants in this generation are hybrids and typically show the dominant traits of both parents combined with heterosis for some characters.
- From F2 onwards, the generations are termed segregating generations and are handled by pedigree, bulk, backcross, or SSD method.
|