PB Ch 22. Breeding in cross Pollinated Crops and Hardy-Weinberg Law
- To understand plant breeding, you first have to understand plant mating habits. Plants generally fall into two categories: self-pollinated (like wheat and rice, which fertilize themselves) and cross-pollinated (like maize, sunflower, onion, and carrots, which rely on wind or insects to mix their pollen with other plants).
- Because cross-pollinated crops constantly swap DNA with their neighbors, they have evolved to thrive on genetic diversity. This fundamental difference requires plant breeders to use entirely different strategies when trying to improve them.
The Challenge: Heterozygosity and Inbreeding Depression
- In nature, cross-pollinated plants are highly heterozygous. This means that for any given trait, the plant carries two different versions of a gene (called alleles)—one inherited from each parent. Every single plant in a field of corn is a unique genetic individual, much like human siblings.
- This diversity is their strength. It creates "hybrid vigor," making the plants tall, resilient, and high-yielding. However, this creates a unique problem for breeders:
- Inbreeding Depression: If a breeder forces a cross-pollinated plant to self-pollinate (mating it with itself to isolate a good trait), the offspring quickly lose their vigor. They become stunted, weak, and lower-yielding. This happens because self-pollination reduces genetic diversity and uncovers hidden, harmful recessive genes.
The Paradigm Shift: Breeding Populations, Not Individuals
- Because of inbreeding depression, a breeder working with cross-pollinated crops cannot simply find one "perfect" plant and clone its seeds. The individual plant's specific genetics (its genotype) are actually of little importance.
- Instead, the breeder's goal is to improve the population as a whole. Think of the crop not as a field of individuals, but as a giant "gene pool." The goal is to increase the frequency of desirable alleles (good genes) in that pool while maintaining enough genetic diversity to keep the plants vigorous.
To achieve this, breeders use specialized methods designed to exploit and maintain this diversity:
- Mass Selection: The simplest method. The breeder visually selects the best plants in a field, harvests their seeds, mixes them together (bulking), and plants them the next year. It improves the average quality of the field without causing inbreeding.
- Population Improvement (Recurrent Selection): A cyclical process of evaluating plants, selecting the best, and intermating them to steadily build up good genes generation after generation.
- Synthetic Varieties: Breeders identify a few excellent, somewhat inbred parent lines that are known to cross well with each other (high General Combining Ability). These are planted together and allowed to open-pollinate, creating a diverse but superior crop.
- Composite Varieties: Similar to synthetics, but created by mixing the seeds of many good-looking lines, regardless of whether their specific cross-compatibility has been tested.
- Hybrid Varieties: The most famous method (especially in corn). Breeders intentionally create highly inbred, weak parent lines to fix specific traits. Then, they cross two distinct inbred lines. The resulting $F_1$ generation (the hybrid) explodes with maximum "heterosis" or hybrid vigor, resulting in uniform, massive yields.
- Clonal Selection: Used for cross-pollinated crops that can be grown from cuttings or roots (like sugarcane). Once a great plant is found, it is simply cloned vegetatively, bypassing seeds entirely.
The Math Behind the Gene Pool: Hardy-Weinberg Law
If breeders are trying to manipulate the "gene pool" of a population, they need a way to measure it. This brings us to the Hardy-Weinberg Law, the absolute foundation of population genetics, established in 1908-1909 by G.H. Hardy and Wilhelm Weinberg.
The Law States: In a large, randomly mating population with no evolutionary forces acting upon it, the frequencies of alleles (genes) and genotypes will remain completely constant from generation to generation.
In other words, if a field of corn is left completely alone, the ratio of good genes to bad genes won't change over time. It sits in a state of equilibrium.
The Mathematical Formulation
Imagine a gene with two versions: a dominant allele ($A$) and a recessive allele ($a$).
- Let p be the frequency of the A allele in the population.
- Let q be the frequency of then a allele in the population.
- Because these are the only two options, their frequencies must add up to 100% (or 1): p + q = 1
- When these plants randomly mate, the frequencies of their offspring's genetic makeup (genotypes) can be predicted by squaring the allele equation (p + q)^2 = 1, which gives us the Hardy-Weinberg equilibrium equation: p^2 + 2pq + q^2 = 1
- p^2 = The fraction of the population that is AA (homozygous dominant)
- 2pq = The fraction that is Aa (heterozygous)
- q^2 = The fraction that is aa (homozygous recessive)
How to calculate it from a real field:
If you have a total number of plants (N), and you count how many are AA (D), Aa (H), and aa (R), you can find the exact allele frequencies using these formulas:
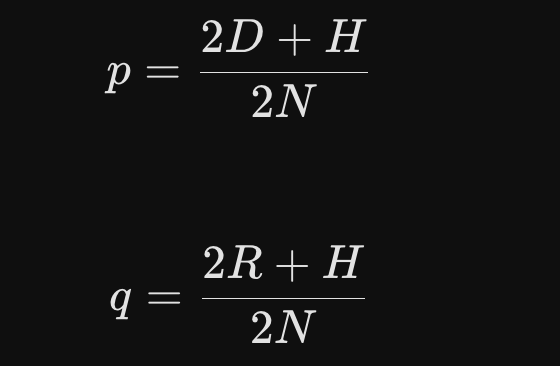
Breaking the Rules: Disturbing the Equilibrium
The Hardy-Weinberg equilibrium describes a population at rest. However, plant breeding is the intentional disruption of this equilibrium. Breeders want to change the allele frequencies to make the crop better.
There are four primary forces that disturb the equilibrium. Breeders use some to their advantage and try to mitigate others:
- Selection: This is the breeder's primary tool. By choosing only the best plants to reproduce, different genotypes have differential reproduction rates. This artificially forces the desirable allele frequency (p) to increase and the undesirable one (q) to decrease.
- Migration: Introducing individuals from an outside population. In breeding, this is done intentionally via intervarietal crosses (bringing in pollen from a different crop variety to introduce brand new alleles).
- Random Genetic Drift: In small populations, allele frequencies can randomly fluctuate due to sheer chance (sampling error). Because breeding programs often use small populations of elite plants, some genetic drift is inevitable.
- Mutation: A random change in the DNA that creates a completely new allele. In nature, this is incredibly slow (about 1 in a million per gene, per generation). Unless a breeder is using directed mutagenesis (like radiation or CRISPR), natural mutation is too slow to be a major factor in standard breeding timelines.
Crucial Distinction: Inbreeding vs. Allele Frequencies
It is vital to understand that inbreeding does NOT change allele frequencies (p and q stay exactly the same). Inbreeding only changes the genotype frequencies. It takes the heterozygous plants (Aa) and splits them into homozygous extremes (AA and aa). The total pool of A and a in the field remains mathematically identical; they are just repackaged into individual plants that now suffer from inbreeding depression.